
Fluoresenssimikroskooppi - Fluorescence microscope


Fluoresenssimikroskoopilla on optisella mikroskoopilla , joka käyttää fluoresenssi sijasta, tai sen lisäksi, sironta , heijastus , ja vaimennus tai imeytymistä , tutkia ominaisuuksia orgaanisia tai epäorgaanisia aineita. "Fluoresenssimikroskooppi" tarkoittaa mitä tahansa mikroskooppia, joka käyttää fluoresenssia kuvan luomiseen, olipa kyseessä yksinkertainen kokoonpano, kuten epifluoresenssimikroskooppi, tai monimutkaisempi rakenne, kuten konfokaalimikroskooppi , joka käyttää optista leikkausta saadakseen paremman resoluution fluoresenssikuvassa .
Periaate
Näyte valaistaan tietyn aallonpituuden (tai aallonpituuksien) valolla, jonka fluoroforit absorboivat , jolloin ne säteilevät pidempien aallonpituuksien valoa (eli eri väriä kuin absorboitu valo). Valaistusvalo erotetaan paljon heikommin säteilevästä fluoresenssista käyttämällä spektrisäteilysuodatinta. Tyypillisiä fluoresenssimikroskoopin komponentteja ovat valonlähde ( ksenonkaarilamppu tai elohopeahöyrylamppu ovat yleisiä; kehittyneemmät muodot ovat suuritehoisia LED - valoja ja lasereita ), virityssuodatin , dikrooninen peili (tai dikrooninen säteensiirto ) ja emissio suodatin (katso alla oleva kuva). Suodattimet ja dikrooninen säteenjakaja valitaan näytteen merkitsemiseen käytetyn fluoroforin spektriherätys- ja päästöominaisuuksien mukaan. Tällä tavalla kuvataan yksittäisen fluoroforin (väri) jakauma kerrallaan. Monivärisiä kuvia useista fluoroforeista on koostettava yhdistämällä useita yksivärisiä kuvia.
Useimmat käytössä olevat fluoresenssimikroskoopit ovat epifluoresenssimikroskooppeja, joissa fluoroforin viritys ja fluoresenssin havaitseminen suoritetaan saman valopolun kautta (eli objektiivin kautta). Näitä mikroskooppeja käytetään laajalti biologiassa ja ne ovat perusta kehittyneemmille mikroskooppisuunnitelmille, kuten konfokaaliselle mikroskoopille ja kokonaisheijastusfluoresenssimikroskoopille (TIRF).
Epifluoresenssimikroskopia

Suurin osa fluoresenssimikroskoopeista, erityisesti biotieteissä käytetyt , ovat kaavion epifluoresenssisuunnittelua. Valossa eksitaatioaallonpituudella valaisee näytteen läpi tavoite linssin. Näytteen lähettämä fluoresenssi kohdistetaan ilmaisimeen samalla objektiivilla, jota käytetään herätykseen ja joka vaatii suurempaa resoluutiota varten objektiivin, jolla on suurempi numeerinen aukko . Koska suurin osa viritysvalosta lähetetään näytteen läpi, vain heijastunut herätysvalo saavuttaa kohteen yhdessä säteilevän valon kanssa ja epifluoresenssimenetelmä antaa siksi korkean signaali-kohinasuhteen. Dikrooninen säteenjakaja toimii aallonpituusspesifisenä suodattimena, joka lähettää fluoresoivaa valoa okulaariin tai ilmaisimeen, mutta heijastaa jäljellä olevan viritysvalon takaisin kohti lähdettä.
Valonlähteet
Fluoresenssimikroskopia vaatii voimakasta, lähes yksiväristä valaistusta, jota jotkut laajalle levinneet valonlähteet, kuten halogeenilamput, eivät pysty tarjoamaan. Neljä pääasiallista valonlähteen käytetään, mukaan lukien ksenonkaarilampusta tai elohopealamppuja , jossa on virityssuodatin , laserit , supercontinuum lähteistä, ja korkea-teho LED . Laserit käytetään laajimmin monimutkaisempien fluoresenssimikroskooppisesti tekniikoita kuten konfokaalimikroskopia ja sisäisen kokonaisheijastuksen fluoresenssimikroskooppisesti taas ksenonlamppu, ja elohopea lamput ja LEDit kanssa dikroinen virityssuodinjärjestelmänä käytetään yleisesti Widefield epifluoresenssin mikroskoopit. Laittamalla kaksi mikrolinssiryhmää laajakenttäisen epifluoresenssimikroskoopin valaistusreitille voidaan saavuttaa erittäin tasainen valaistus, jonka vaihtelukerroin on 1-2%.
näytteen valmistus
Näyttää peittokuvat neljästä fluoresoivasta kanavasta
(b) Syaani: [PLL -A546 -fluoresenssi] - yleinen vastaväri eukaryoottisten solupintojen visualisoimiseksi
(c) Sininen: [Hoechst -fluoresenssi] - värjää DNA: n, tunnistaa ytimet
(d) Punainen: [klorofyllin autofluoresenssi] - erottaa kloroplastit

Jotta näyte soveltuisi fluoresenssimikroskopiaan, sen on oltava fluoresoiva. Fluoresoivan näytteen luomiseen on useita menetelmiä; tärkeimmät tekniikat ovat leimaamalla fluoresoivilla tahroja tai, jos kyseessä on biologisten näytteiden, ilmentyminen on fluoresoiva proteiini . Vaihtoehtoisesti voidaan käyttää näytteen luontaista fluoresenssia (eli autofluoresenssia ). Biotieteissä fluoresenssimikroskopia on tehokas työkalu, joka mahdollistaa näytteen spesifisen ja herkän värjäyksen proteiinien tai muiden kiinnostavien molekyylien jakautumisen havaitsemiseksi . Tämän seurauksena biologisten näytteiden fluoresoivalla värjäyksellä on erilaisia tekniikoita.
Biologiset fluoresoivat tahrat
Monet fluoresoivat tahrat on suunniteltu useille biologisille molekyyleille. Jotkut näistä ovat pieniä molekyylejä, jotka ovat luontaisesti fluoresoivia ja sitovat kiinnostavaa biologista molekyyliä. Tärkeimpiä esimerkkejä näistä ovat nukleiinihappovärjäykset , kuten DAPI ja Hoechst (kiihdytetään UV -aallonpituuden valolla) ja DRAQ5 ja DRAQ7 (optimaalisesti punaisen valon herättämä), jotka kaikki sitovat DNA: n pienen uran ja merkitsevät siten solujen ytimet . Muut ovat lääkkeitä, toksiineja tai peptidejä, jotka sitovat tiettyjä solurakenteita ja jotka on johdettu fluoresoivalla reportterilla. Merkittävä esimerkki tämän luokan fluoresoiva tahra on -falloidiinia , jota käytetään värjäämään aktiinin kuituja nisäkkäiden soluissa. Uusi peptidi, joka tunnetaan nimellä Collagen Hybridizing Peptide , voidaan myös konjugoida fluoroforien kanssa ja käyttää denaturoitujen kollageenikuitujen värjäämiseen . Kasvisoluseinien värjäys suoritetaan käyttämällä tahroja tai väriaineita, jotka sitovat selluloosaa tai pektiiniä . Pyrkimys fluoresoiviin koettimiin, joilla on suuri spesifisyys, mahdollistavat myös kasvisolujen elävän kuvantamisen, on käynnissä.
On olemassa monia fluoresoivia molekyylejä, joita kutsutaan fluoroforeiksi tai fluorokromeiksi , kuten fluoreskeiini , Alexa Fluors tai DyLight 488 , jotka voidaan yhdistää kemiallisesti toiseen molekyyliin, joka sitoo kiinnostavan kohteen näytteessä.
Immunofluoresenssi
Immunofluoresenssi on tekniikka, jossa käytetään vasta -aineen erittäin spesifistä sitoutumista sen antigeeniin spesifisten proteiinien tai muiden solussa olevien molekyylien leimaamiseksi. Näyte käsitellään ensisijaisella vasta -aineella, joka on spesifinen kiinnostuksen kohteena olevalle molekyylille. Fluorofori voidaan konjugoida suoraan primaariseen vasta -aineeseen. Vaihtoehtoisesti voidaan käyttää sekundaarista vasta -ainetta, joka on konjugoitu fluoroforiin ja joka sitoutuu spesifisesti ensimmäiseen vasta -aineeseen. Esimerkiksi hiirissä kasvatettua primaarista vasta-ainetta, joka tunnistaa tubuliinin yhdistettynä toissijaiseen hiiren vastaiseen vasta-aineeseen, joka on johdettu fluoroforilla, voitaisiin käyttää solun mikrotubulusten leimaamiseen .
Fluoresoivat proteiinit
Nykyaikainen käsitys genetiikasta ja DNA: n muokkaamiseen käytettävissä olevista tekniikoista antaa tutkijoille mahdollisuuden muuntaa geneettisesti proteiineja kuljettamaan myös fluoresoivan proteiinin reportteria. Biologisissa näytteissä tämä sallii tutkijan tehdä suoraan kiinnostavan proteiinin fluoresoivaksi. Proteiinin sijaintia voidaan sitten seurata suoraan, myös elävissä soluissa.
Rajoitukset
Fluoroforit menettävät kykynsä fluoresoida, kun ne valaistuvat prosessissa, jota kutsutaan valonvalkaisuksi . Valovalkaisu tapahtuu, kun fluoresoivat molekyylit keräävät kemiallisia vaurioita fluoresenssin aikana herätetyistä elektroneista. Valovalkaisu voi rajoittaa voimakkaasti aikaa, jonka kuluessa näyte voidaan havaita fluoresenssimikroskoopilla. Olemassa useita tekniikoita vähentää valovalkaisu kuten käyttämällä järeämmästä fluoroforin, minimoimalla valaistus, tai käyttämällä valolta scavenger kemikaaleja.
Fluoresenssimikroskopia fluoresoivilla reportteriproteiineilla on mahdollistanut elävien solujen analysoinnin fluoresenssimikroskoopilla, mutta solut ovat alttiita fototoksisuudelle, erityisesti lyhyen aallonpituuden valossa. Lisäksi fluoresoivilla molekyyleillä on taipumus synnyttää reaktiivisia kemiallisia lajeja valaistessaan, mikä parantaa fototoksista vaikutusta.
Toisin kuin läpäisseet ja heijastuneet valomikroskopiatekniikat, fluoresenssimikroskopia sallii vain tiettyjen rakenteiden havaitsemisen, jotka on merkitty fluoresenssiksi. Esimerkiksi fluoresoivalla DNA -värillä valmistetun kudosnäytteen tarkkailu fluoresenssimikroskoopilla paljastaa vain DNA: n organisaation soluissa eikä paljasta mitään muuta solun morfologioista.
Laskentatekniikat, joilla ehdotetaan fluoresoivan signaalin arviointia ei-fluoresoivista kuvista (kuten kirkaskenttä), voivat vähentää näitä huolenaiheita. Yleensä näihin lähestymistapoihin kuuluu syvän konvoluution hermoverkoston kouluttaminen värjättyihin soluihin ja sitten fluoresenssin arvioiminen värjäämättömissä näytteissä. Siten irrottamalla tutkittavat solut verkon kouluttamiseen käytetyistä soluista, kuvantaminen voidaan suorittaa nopeammin ja pienemmällä valotoksisuudella.
Subdiffraktiotekniikat
Valon aallon luonne rajoittaa sen pisteen kokoa, johon valo voidaan keskittyä diffraktiorajan vuoksi . Ernst Abbe kuvasi tätä rajoitusta 1800 -luvulla ja "rajoittaa optisen mikroskoopin resoluution noin puoleen käytetyn valon aallonpituudesta". Fluoresenssimikroskopia on keskeinen monissa tekniikoissa, joiden tavoitteena on ylittää tämä raja erikoisoptiikkakokoonpanoilla.
1900 -luvulla on keksitty useita parannuksia mikroskooppitekniikoihin, ja ne ovat johtaneet jossain määrin parempaan resoluutioon ja kontrastiin. He eivät kuitenkaan ylittäneet diffraktiorajaa. Vuonna 1978 kehitettiin ensimmäiset teoreettiset ajatukset tämän esteen rikkomiseksi käyttämällä 4Pi-mikroskooppia konfokaalisena laserskannausfluoresenssimikroskooppina, jossa valo kohdistetaan ihanteellisesti kaikilta puolilta yhteiseen tarkennukseen, jota käytetään kohteen skannaamiseen piste-kohdalta piste-heräte yhdistettynä piste-piste-havaintoon. Ensimmäinen kokeellinen 4pi-mikroskoopin esittely tapahtui kuitenkin vuonna 1994. 4Pi-mikroskooppi maksimoi käytettävissä olevien tarkennussuuntien määrän käyttämällä kahta vastakkaista objektiivia tai kahden fotonin viritysmikroskopiaa käyttämällä punasiirtynyttä valoa ja monifotoniviritystä.
Integroitu korrelatiivinen mikroskooppi yhdistää fluoresenssimikroskoopin elektronimikroskoopin kanssa. Tämä mahdollistaa ultrastruktuurin ja asiayhteyteen liittyvien tietojen visualisoinnin elektronimikroskoopilla samalla kun käytetään fluoresenssimikroskoopin tietoja etiketöintityökaluna.
Ensimmäinen tekniikka subdiffraktiotarkkuuden saavuttamiseksi oli STED-mikroskooppi , jota ehdotettiin vuonna 1994. Tämä menetelmä ja kaikki RESOLFT- konseptia seuraavat tekniikat perustuvat voimakkaaseen epälineaariseen vuorovaikutukseen valon ja fluoresoivien molekyylien välillä. Molekyylejä ohjataan voimakkaasti erotettavissa olevien molekyylitilojen välillä kussakin tietyssä paikassa, joten lopulta valoa voidaan lähettää vain pienessä osassa tilaa, mikä lisää resoluutiota.
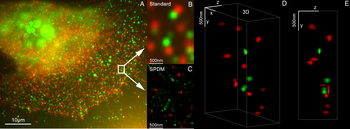
Samoin 1990 -luvulla on kehitetty toinen laajakenttämikroskopiaan perustuva superresoluutiomikroskopiamenetelmä. Fluoresoivalla markkerilla värjättyjen solunanorakenteiden koon erottuminen merkittävästi parannettiin kehittämällä SPDM -paikannusmikroskopia ja strukturoitu laservalaistus (spatiaalisesti moduloitu valaistus, SMI). SPDM -periaatteen ja SMI: n yhdistäminen johti Vertico SMI -mikroskoopin kehittämiseen. Normaalien vilkkuvien fluoresoivien väriaineiden, kuten vihreän fluoresoivan proteiinin (GFP), yksittäismolekyylinen havaitseminen voidaan saavuttaa käyttämällä SPDM: n edelleen kehittämistä, niin sanottua SPDMphymod-tekniikkaa, joka mahdollistaa kahden erilaisen fluoresoivan molekyylin havaitsemisen ja laskemisen molekyylitasolla (tämä tekniikkaa kutsutaan kaksiväriseksi lokalisointimikroskopiaksi tai 2CLM: ksi).
Vaihtoehtoisesti fotoaktivoidun lokalisointimikroskopian tulo voisi saada samanlaisia tuloksia luottaen yksittäisten molekyylien vilkkumiseen tai vaihtamiseen, jolloin fluoresoivien molekyylien osuus on hyvin pieni joka kerta. Tämä molekyylien stokastinen vaste levitetylle valolle vastaa myös erittäin epälineaarista vuorovaikutusta, joka johtaa subdiffraktion resoluutioon.
Fluoresenssimikrokuvagalleria

Osteosarkoomasolun z-projektio, joka on värjätty phalloidiinilla aktiinifilamenttien visualisoimiseksi. Kuva otettiin konfokaalisella mikroskoopilla, ja seuraava dekonvoluutio tehtiin käyttämällä kokeellisesti johdettua pisteen hajautusfunktiota.
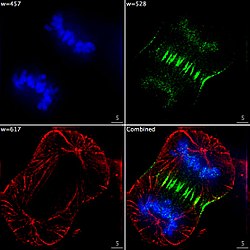
Epifluoresenssikuvaus kolmesta komponentista jakautuvassa ihmisen syöpäsolussa. DNA värjätään sininen, joka on proteiini nimeltään INCENP on vihreä, ja mikrotubulusten on punainen. Kukin fluorofori kuvataan erikseen käyttäen eri herätys- ja päästösuodattimien yhdistelmää, ja kuvat otetaan peräkkäin käyttämällä digitaalista CCD -kameraa , minkä jälkeen ne peitetään täydellisen kuvan saamiseksi.

Endoteelisolut mikroskoopin alla. Ytimet on värjätty siniseksi DAPI: llä, mikrotubulukset on merkitty vihreällä FITC: hen sitoutuneella vasta -aineella ja aktiinifilamentit on merkitty punaiseksi ja TRITC: hen sitoutunut falloidiini. Naudan keuhkovaltimon endoteelisolut (BPAE)
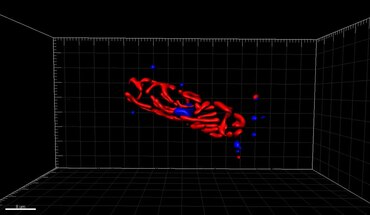
3D-kaksivärinen superresoluutiomikroskopia, jossa Her2 ja Her3 rintasoluissa, vakiovärit: Alexa 488, Alexa 568. LIMON-mikroskooppi

Ihmisen lymfosyyttien ydin, joka on värjätty DAPI: llä ja kromosomi 13 (vihreä) ja 21 (punainen) sentromeerikoettimet hybridisoituneet ( fluoresoiva in situ -hybridisaatio (FISH))

Hiivasolukalvo visualisoitu joidenkin RFP- ja GFP -fluoresoivien merkkiaineiden kanssa fuusioituneiden kalvoproteiinien avulla. Molempien merkkien valon tuottaminen johtaa keltaiseen väriin.

Supertarkkuusmikroskopia: YFP-molekyylin havaitseminen ihmisen syöpäsolussa. Tyypilliset etäisyysmittaukset 15 nm alueella mitattuna Vertico-SMI/SPDMphymod-mikroskoopilla

Supertarkkuusmikroskopia: Paikallismikroskopia (2CLM) GFP- ja RFP-fuusioproteiinien (luusyöpäsolun ydin) kanssa 120 000 lokalisoitua molekyyliä laajakenttäalueella (470 µm 2 ) mitattuna Vertico-SMI/SPDMphymod-mikroskoopilla

Fluoresenssimikroskopia DNA-ilmentymisestä ihmisen villityypissä ja P239S-mutantti- Palladinissa .

Fluoresenssimikroskooppikuvat auringonsäteilyn patologiasta verisolussa, jossa vaikutusalueet näkyvät punaisina.










Katso myös
- Fluoresenssikuvaus
- Fluoresenssi biotieteissä
- Korrelatiivinen valo-elektronimikroskopia
- Elizabeth Harry , edelläkävijä fluoresenssimikroskopiatekniikoissa bakteerien subcellulaaristen proteiinien visualisoimiseksi
- Vihreä fluoresoiva proteiini (GFP)
- Elohopeahöyrylamppu
- Mikroskooppi
- Skannaava elektronimikroskooppi#katodoluminesenssi
- Stokesin muutos
- Xenon -valolamppu
Viitteet
Ulkoiset linkit
|
Kirjastoresursseja noin Fluoresenssimikroskopia |
- Fluorophores.org , fluoresoivien väriaineiden tietokanta
- Mikroskopian resurssikeskus
- animaatioita ja selityksiä erityyppisistä mikroskoopeista, mukaan lukien fluoresoivat ja konfokaliset mikroskoopit (Université Paris Sud)