
Dinosaur Park -muodostus - Dinosaur Park Formation
| Dinosaur Park Formation Stratigraphic -alue : myöhäinen liitu , Campanian ,76,9-75,8 Ma | |
|---|---|
 Dinosaur Park Formation altistuvat pitkin Red Deer River in Dinosaur Provincial Park , Kaakkois Kanadan Albertassa.
| |
| Tyyppi | Geologinen muodostuminen |
| Yksikkö | Belly River Group |
| Pohjana | Karhunpahvin muodostus |
| Päälliset | Oldmanin muodostus |
| Litologia | |
| Ensisijainen |
Hiekkakivi (alempi) mudstone ja siltstone (ylempi) |
| Muut | Bentoniitti ja hiili |
| Sijainti | |
| Koordinaatit | 49 ° 12'N 110 ° 24'W / 49,2 ° N 110,4 ° W Koordinaatit : 49,2 ° N 110,4 ° W49 ° 12'N 110 ° 24'W / |
| Arvioidut paleokoordinaatit | 56 ° 24'N 75 ° 48'W / 56,4 ° N 75,8 ° W |
| Alue |
|
| Maa |
|
| Laajuus | Länsi -Kanadan sedimenttiallas |
| Tyyppi -osio | |
| Nimetty | Dinosaur Provincial Park |
| Nimetty | Eberth, DA ja Hamblin, AP |
| Vuosi määritelty | 1993 |
 Dinosaur Park Formation (Kanada)
 Dinosaur Park Formation (Alberta)
| |
Dinosaur Park muodostuminen on ylin jäsen vatsa-joen ryhmän (tunnetaan myös Judith River ryhmän ), joka on merkittävä geologinen yksikkö Etelä Alberta . Se talletettiin myöhäisen liitukauden Campanian vaiheen aikana , noin 76,9-75,8 miljoonaa vuotta sitten. Se on talletettu tulva ja rannikon tavallinen ympäristöissä, ja se rajoittuu nonmarine Oldman muodostaminen alle sen ja meren Bearpaw muodostaminen sen yläpuolella.
Dinosaur Park Formation sisältää tiheitä dinosaurusluurankoja , sekä nivellettyjä että disartikuloituja, joita esiintyy usein pehmytkudosten säilyneiden jäännösten kanssa. Myös muiden eläinten, kuten kalojen , kilpikonnien ja krokotiilien , jäänteitä sekä kasvien jäänteitä on runsaasti. Muodostuma on nimetty Unescon maailmanperintökohteisiin kuuluvan Dinosaur Provincial Park -puiston mukaan , jossa muodostuma on hyvin paljastettu Red Deer -joen reunalla olevilla badlandsilla .
Geologinen asetus

Dinosaur Park Formation koostuu sedimentteistä, jotka ovat peräisin vuorten eroosiosta länteen. Se laskeutui tulva -rannikkotasangolle jokijärjestelmillä, jotka virtaavat itään ja kaakkoon Bearpaw -merelle, suurelle sisämerelle, joka oli osa Länsi -sisäistä meritietä . Että meri vähitellen hukkua viereisen rannikon plain, kerrostamalla meren savikiven että Bearpaw muodostaminen päälle Dinosaur Park Formation.
Dinosaur Park Formation on noin 70 metriä (230 jalkaa) paksu Dinosaur Parkissa. Muodostelman alempi osa sijoitettiin virtauskanavaympäristöihin ja koostuu pääasiassa hienoista tai keskijyväisistä , ristikkäisistä hiekkakivistä . Ylemmän osan, joka on talletettu overbank ja tulva ympäristöissä, koostuu pääasiassa massiivinen ja laminoitu, orgaaninen-rikas mudstones runsaasti juuri jälkiä, ja ohut sängyt bentoniitin . Lethbridgen hiilivyöhyke, joka koostuu useista matalan asteen hiilen saumoista, jotka on yhdistetty muta- ja kivikiviin , merkitsee muodostuman huippua.
Dinosaur Park Formationin sedimentit ovat samanlaisia kuin taustalla olevan Oldman Formationin sedimentit, ja ne sisältyivät alun perin tähän muodostumiseen. Näitä kahta muodostumista erottaa kuitenkin alueellinen ristiriita , ja ne erottuvat petrografisista ja sedimentologisista eroista. Lisäksi nivelletyt luuston jäänteet ja luut ovat harvinaisia Oldmanin muodostumassa, mutta runsaasti Dinosauruspuiston muodostumassa.
Biostratigrafia
Dinosaur Park Formation voidaan jakaa ainakin kahteen erilliseen eläimistöön. Muodostelman alaosalle on ominaista Corythosauruksen ja Centrosauruksen runsaus . Tämä lajiryhmä korvataan muodostumassaan korkeammalla eri ornithischian eläimistölle, jolle on ominaista Lambeosaurus ja Styracosaurus . Useiden uusien, harvinaisten ornithischia- lajien esiintyminen muodostuman huipulla voi osoittaa, että kolmas erillinen eläimistö oli korvannut toisen siirtyessään nuorempiin, ei-dinosauruspuiston sedimentteihin, samalla kun sisämeri siirtyy maahan mutta täällä on vähemmän jäänteitä. Nimetön pachyrhinosaurus , Vagaceratops irvinensis ja Lambeosaurus magnicristatus voivat olla yleisempiä tässä kolmannessa eläimistössä.
Alla oleva aikajana seuraa Fowlerin (2016) esittämää synteesiä, jossa on lisätietoja Arbor et al. 2009, Evans et ai. 2009 ja Penkalski, 2013. Megaherbivore Assemblage Zones (MAZ) seuraavat Mallon et al. , 2012.

Sammakkoeläimet
Muodostelmasta on löydetty seuraavien sammakkoeläinten jäännöksiä:
Albanerpetontidae (sukupuuttoon kuolleet, salamanderimaiset sammakkoeläimet)
- Habrosaurus prodilatus
- Lisserpeton
- Opisthotriton kayi
- Scapherpeton tectum
- nimetön caudatan
- 2 määrittelemätöntä caudatania
- 2 nimeämätöntä salientania
- Tyrrellbatrachus brinkmani
- Hensonbatrachus kermiti
Dinosaurukset
Muodostelmasta on löydetty seuraavien dinosaurusten jäännöksiä:
Ornithischians
Muodostumasta on löydetty seuraavien ornitistien jäänteitä:
Ankylosaurs
| Ankylosaurukset Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| Anodontosaurus | A. inceptus | Keskellä, 75,6 vuotta sitten | [Kaksi] kalloa hampailla, alaleuka, osittain valmis luuranko, molemmat kohdunkaulan puolirenkaat ja osteodermit. | Ankylosaurine ankylosaurid |
 |
|
| Dyoplosaurus | D. acutosquameus | Alempi, 76,5 vuotta sitten | Osittainen kallo ja luuranko, mukaan lukien lantion, hännän ja takaraajan pesillä ja osteodermillä. | Ankylosaurine ankylosaurid |
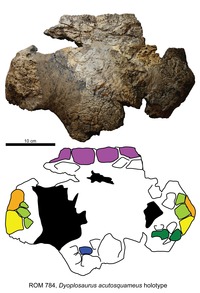 |
|
| Edmontonia | E. rugosidens | Alempi, 76,5-75,9 Ma sitten | Osittainen luuranko, joka sisältää kallon, selkärangan, proksimaalisen, distaalisen kaudaalisen, kylkiluut, olkaluun, kyynärpään, säteen, manuksen, lantion palaset, sääriluun, fibulan?, Osteodermat; nivelletun etupuolen, jossa on in situ osteodermit ja pariksi muodostetut ensimmäiset mediaalit. | Nodosaurine nodosaurid tunnetaan myös Horseshoe Canyon muodostaminen ja kaksi lääkettä Formation |
 |
|
| Euoplocephalus | E. tutus | Alemmasta keskelle, ~ 76.4-75.6 Ma sitten | [Neljä] kalloa, alalehteä, kohdunkaulan nikamia, selkärankaa, kylkiluita, lapaluita, olkaluu, säde, kyynärpää, käsivarren rintakehä, phalanx, ilium, ischium, reisiluu, sääriluu, osittainen pes, ristiluu, kohdunkaulan puolirenkaat ja osteodermit. | Ankylosaurine ankylosaurid |
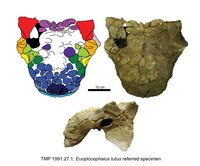 |
|
| Panoplosaurus | P. mirus | Keskellä, 75,6 vuotta sitten | Kallo, jossa on alaleuat, eristetyt hampaat, kohdunkaulan nikamat, selkärangan, ristiluun nikamat, kohdunkaulan kylkiluut, dorsaaliset kylkiluut, scapulocoracoid, olkaluu, manus, sääriluu, fibula, luutunut intersternal levy, pari luutuneita xiphoid -prosesseja, pes ja in situ osteodermit . | Nodosaurine nodosaurid |
 |
|
| Platypelta | P. coombsi | Alempi, 77,5-76,5 Ma sitten | Hyvin säilynyt kallo, alaleuat, hampaat, kohdunkaulan ja selkärangan, kylkiluut, täydellinen lantio, molemmat scapulocoracoids, sekä humeri että säteet, molemmat kohdunkaulan puolirenkaat ja osteodermit. | Ankylosaurine ankylosaurid |
 |
|
| Scolosaurus | S. cutleri | Alempi, 76,5 miljoonaa vuotta sitten tai enemmän | Lähes täydellinen luuranko, kallo, kohdunkaula-, selkä- ja niska-nikamat, kylkiluut, lapaluut, coracoid, humeri, säteet, ilium, ischium, reisiluut, sääriluu, fibula, [yksi] kohdunkaulan puolirengas ja osteodermit. | Ankylosaurine ankylosaurid ajateltiin lyhyesti olevan Euoplocephalus -synonyymi . Se on mahdollisesti peräisin taustalla olevan Oldman -muodostuman ylemmistä kerroksista . |
 |
|
| S. thronus | Ylempi, 75 vuotta sitten | Osittainen luuranko, joka sisältää kallon, selkärangan,? Täydellisen synsacrumin, ristiluun kylkiluut, kaulan nikamat, lapaluun, osittaisen lonkan, olkaluun, kohdunkaulan puolirenkaat, osteodermit ja ihon jäljet. | Ankylosaurine ankylosaurid | |||
Ceratopsians
Nimeämätön Pachyrhinosaurus -kaltainen taksoni on saatu talteen muodostumasta.
| Ceratopsians Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| Centrosaurus | C. apertus | Keskellä, 76.2-75.5Ma sitten | "[Viisitoista] kalloa, useita luurankoja, kaikki aikuisia; runsaasti luustoa sisältävää materiaalia, jossa on harvinaisia nuoria ja alaikäisiä." C. nasicornis voi olla synonyymi. | Centrosaurine ceratopsid |
 |
|
| Chasmosaurus | C. belli | Keskellä, 76-75.5Ma sitten | "[Kaksitoista] kalloa, useita luurankoja." | Chasmosaurine ceratopsid |
 |
|
| C. russelli | Alempi, 76,5-76Ma sitten | "[Kuusi] täydellistä tai osittaista pääkalloa." | ||||
| Mercuriceratops | M. gemini | Alempi, ~ 77Ma sitten | "yksi apomorfinen squamosal" | Chasmosaurine ceratopsid |
 |
|
| Monoklonius | M. lowei | Epäilyttävä centrosaurine ceratopsid. Mahdollisesti synonyymi Centrosaurukselle . | ||||
| Pentaceratops | P. aquilonius | Ylin, 74,8 MA | kaksi rypytystä | Epäilyttävä chasmosaurine ceratopsid, joka voi olla sama laji kuin Spiclypeus shipporum . |
 |
|
| Spinops | S. sternbergorum | Alempi, 76,5Ma | "osittainen parietaaliluu, osittainen hammas, tunnistamattomat raajafragmentit, osittainen kallo ja osittainen oikea squamosal." | Centrosaurine ceratopsid, joka voi itse asiassa olla ylemmästä Oldman -muodostumasta. |
 |
|
| Styracosaurus | S. albertensis | Ylempi, 75,5-75,2Ma sitten | "[Kaksi] kalloa, [kolme] luurankoa, lisämateriaalia luusängyissä." | Centrosaurine ceratopsid |
 |
|
| Unescoceratops | U. koppelhusae | Osittainen alaleuka | Leptoceratopsid arvellaan olleen vuodesta kahteen metriä pitkä ja alle 91 kiloa. Sen hampaat olivat pyöreimmät kaikista leptoceratopsideista. | |||
| Vagaceratops | V. irvinensis | Ylempi, 75Ma sitten | "[Kolme] pääkalloa, luuranko puuttuu hännästä." | Chasmosaurine ceratopsid laji on aikaisemmin luokiteltu lajiksi Chasmosaurus . |
 |
|
Ornitopodit
Ainakin yksi määrittämätön hypsilophodont -näyte on otettu talteen.
Darren H. Tanke ja MK Brett-Surman päättivät vuonna 2001 tehdyssä katsauksessa hadrosauruksen munankuorista ja kuoriutumismateriaalista Dinosaur Park Formationista, että hadrosaurukset pesivät sekä muinaisella ylängöllä että muodostuman laskeumaympäristön alamailla. Harvemmat hadrosaurukset, kuten Brachylophosaurus tai Parasaurolophus, ovat saattaneet suosia ylängön pesimäpaikkoja . Kirjoittajat eivät kuitenkaan pystyneet määrittämään, mitkä erityiset tekijät vaikuttivat pesimäpaikan valintaan muodostelman hadrosauruksissa. He ehdottivat, että käyttäytyminen, ruokavalio, maaperän kunto ja kilpailu dinosauruslajien välillä vaikuttivat potentiaalisesti siihen, missä hadrosaurukset pesivät.
Dinosauruspuiston muodostumasta on raportoitu alle senttimetrin palasia pikkukivirakenteista hadrosauruksen munankuorta. Tämä munankuori on samanlainen kuin Devil's Couleen Hadrosaur -munankuori Etelä -Albertassa sekä Two Medicine ja Judith River Formations Montanassa, Yhdysvalloissa. Vaikka dinosaurusten munankuori on läsnä, se on hyvin harvinainen Dinosaur Park Formationissa, ja sitä esiintyy vain kahdessa eri mikrofossiilikohdassa. Nämä sivustot ovat ominaisia suuri määrä pisidiid simpukat ja muut harvinaisempia kuoritut selkärangattomien kuten unionid simpukat ja etanat. Tämä yhteys ei ole sattumaa, koska selkärangattomat kuoret olisivat hitaasti liuennut ja vapauttaneet tarpeeksi emäksistä kalsiumkarbonaattia suojaamaan munankuoria luonnossa esiintyviltä hapoilta, jotka muuten olisivat liuottaneet ne ja estäneet fossiiloitumisen.
Toisin kuin munankuoren fossiilit, hyvin nuorten hadrosaurusten jäänteet ovat itse asiassa hieman yleisiä. Darren Tanke on havainnut, että kokenut keräilijä voisi todella löytää useita nuoria hadrosaurinäytteitä yhdessä päivässä. Yleisimpiä nuorten hadrosaurusten jäänteitä Dinosaur Park Formationissa ovat hampaat , raajojen ja jalkojen luut sekä selkärangan keskusta . Materiaalissa ei näkynyt juurikaan tai lainkaan kulumista, joka olisi aiheutunut kuljetuksesta, mikä tarkoittaa, että fossiilit haudattiin lähelle lähtöpaikkaa. Luunpohjat 23, 28, 47 ja 50 ovat muodostuneiden nuorten hadrosauruksen jäännösten tuottavia lähteitä, etenkin luustoa 50. Nuorten hadrosaurusten ja fossiilisten munankuorifragmenttien luiden ei tiedetä säilyneen yhdessä, vaikka molemmat ovat läsnä muodostelma.
| Ornitopodit Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| Corythosaurus | C. casuarius | Alempi-keskimmäinen, 76,5-75,5Ma sitten | "Noin [kymmenen] nivelkalloa ja niihin liittyvää postkraniaa, [kymmenen -viisitoista] nivelkalloa, eristettyjä kalloelementtejä, nuorista aikuisiin." | Lambeosaurin lambeosaurine hadrosaur |
 |
|
| Gryposaurus | G. notabilis | Alempi, 76,2-76Ma sitten | "Noin [kymmenen] kokonaista pääkalloa, [kaksitoista] sirpaleista pääkalloa, niihin liittyvä postkrania." | Kritosaurin saurolophine hadrosaur |
 |
|
| Lambeosaurus | L. lambei | Ylempi, 75,5-75Ma sitten | "Noin [seitsemän] nivelkalloa, joihin liittyy postkraniaa, [mahdollisesti kymmenen] nivelkalloa, yksittäisiä kalloelementtejä, nuorista aikuisiin." | |||
| L. magnicristatus | Ylä-/ Karhunpaimen muodostus , 74.8Ma sitten | "[Kaksi] täydellistä pääkalloa, joista toiselle liittyy niveltynyt postkrania." |
 |
|||
| Parasaurolophus | P. walkeri | Alempi, 76,5-75,3Ma sitten | "Täydellinen kallo ja postkraniaalinen luuranko." | Parasaurolophin lambeosaurine hadrosaur. |
 |
|
| Prosaurolophus | P. maximus | Ylempi, 75,5 - 74,8 Ma | "[Kaksikymmentä-kaksikymmentäviisi] yksilöä, mukaan lukien vähintään [seitsemän] nivelkalloa ja niihin liittyvä postkrania." | Saurolophin saurolophine hadrosaur |
 |
|
Pachycephalosaurs
| Pachycephalosaurs Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| Foraminacephale | F. brevis | Läsnä myös Oldman -kokoonpanossa | Frontoparetal -kupoli, useita muita kallonpaloja, mukaan lukien nuorten ja subadult -materiaalit | Kun sitä pidettiin Stegoceras -lajina |

|
|
| Gravitholus | G. albertae | "Frontoparietal -kupoli." |
 |
|||
| Hanssuesia | H. sternbergi | Alempi, läsnä myös Oldmanin ja Judith Riverin muodostumissa |
 |
|||
| Stegoceras | S. validum | Näytteet, mukaan lukien frontoparietaalinen kupoli. |
 |
|||
Teropodit
Dinosauruspuiston muodostumassa pienet teropodit ovat harvinaisia, koska niiden ohutseinäisten luiden taipumus murtua tai heikosti säilyttää. Pienten teropodien pienet luut, joita suuret saalistivat, on voitu niellä kokonaisina ja sulattaa. Tässä yhteydessä erityisen arvokas oli löytää pieni teropodinen dinosaurus, jossa oli säilyneitä hammasjälkiä. Mahdolliset määrittelemättömät avimimidijäännökset tunnetaan muodostumasta.
Ornitomimidit
| Ornitomimidit Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| Ornitomimus | O. sp. | Tyyppinäyte | Ornithomimid , mahdollisesti lajin struthiomimus . |
 |
||
| Qiupalong | Q. sp. | Useita yksilöitä | Ornithomimid mahdollisesti säteilyn tämän suvun Aasiaan. |
 |
||
| Ratifioi | R. evadens | Tyyppinäyte | Ornitomimidi, aiemmin Struthiomimuksen näyte . |
 |
||
Oviraptorosaurukset
Värinäppäin
|
Huomautuksia Epävarmat tai alustavat taksonit ovat pienessä tekstissä ; |
| Oviraptorosaurukset Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| Caenagnathus | C. collinsi | Alaleuka, tyyppinen näyte | Caenagnathid joka kilpailivat Anzu koko. |
 |
||
| Chirostenotes | C. pergracilis | Useita sirpaleisia näytteitä, tyyppinen näyte | Keskikokoinen caenagnathid. |
 |
||
| Citipes | C. elegans | Useita sirpaleisia näytteitä, tyyppinen näyte | Pienin caenagnathid muodostumasta. |
 |
||
| Makrofalangia | M. canadensis | Chirostenotes pergracilisin nuoriso synonyymi | ||||
Paravilaiset
Muodostelman yläosasta tunnetaan uusi troodontidin taksoni, joka perustuu yksinomaan hampaisiin.
| Paravians Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| vrt. Baptornis | Määrittämätön | Hesperornithine lintu | ||||
| vrt. Cimolopteryx | Määrittämätön | Osittainen korakoidi | Mahdollinen charadriiform lintu | |||
| Dromaeosaurus | D. albertensis | Useita näytteitä ja hampaita, tyyppinen näyte | Dromaeosaurid |
 |
||
| Hesperonychus | H. elizabethae | Lonkan luut ja osittaiset varpaat ja kynnet, tyyppinen näyte | Microraptorine dromaeosaur, löytyy myös Oldman Formation |
 |
||
| Latenivenatrix | L. mcmasterae | Lonkan luut, lantio, kallon sirpaleet, tyyppinäyte | Suuri troodontidi, jonka pituus on 3–3,5 m (9,8–11,5 jalkaa). |
.png) |
||
| vrt. Palintropus | Nimetön | Osittain olkahihnat | Ambiortiform lintu | |||
| vrt. Paronychodon | vrt. P. lacustris | Hampaat | Määrittämätön maniraptoraani, löytyy myös Judith -joesta | |||
| vrt. Pektiini | Määrittämätön | Hampaat | Troodontti | |||
| Polyodontosaurus | P. grandis | Hampaallinen, tyyppinen näyte | Nimi dubium. Mahdollisesti synonyymi Latenivenatrixille. | |||
| Richardoestesia | R. gilmorei | Alaleuka , tyyppinen näyte | Dromaeosaurid | |||
| R. tasakylkinen | Hampaat | |||||
| Saurornitholestes | S. langstoni | Epätäydellinen luuranko ja hampaat, tyyppinäyte . Dentary tarkoitetut Saurornitholestes havaittiin, että säilynyt hampaan jäljet nuoren tyrannosaur. | Dromaeosaurid |
 |
||
| Stenonychosaurus | S. inequalis | Lähes täydellinen luuranko ja muut osittaiset luurangot, tyyppinen näyte | Troodontid kerran ajateltiin olevan laji Troodon |
 |
||
Tyrannosaurukset
| Tyrannosaurukset Dinosaur Park Formationista | ||||||
|---|---|---|---|---|---|---|
| Suku | Laji | Sijainti | Stratigrafinen sijainti | Materiaali | Huomautuksia | Kuvat |
| Daspletosaurus | Nimeämätön laji | Keskimmäinen, 75,6-75Ma sitten | Useita yksilöitä | Tyrannosaurine tyrannosaurid , myös läsnä Bearpaw Formation |

|
|
| Gorgosaurus | G. libratus | Alempi-keskimmäinen, 76,6-75,1Ma sitten | Lukuisia näytteitä, tyyppinäytteitä | Albertosaurine tyrannosaurid, esiintyvät myös Judith joen muodostaminen ja mahdollisesti kaksi lääkettä muodostaminen . |
 |
|
Värinäppäin
|
Huomautuksia Epävarmat tai alustavat taksonit ovat pienessä tekstissä ; |
Muut matelijat
Kuorodiodit
Choristoderes tai champsosaurs olivat vedessä eläviä matelijoita. Pienet esimerkit näyttivät liskoilta, kun taas suuret tyypit olivat pinnallisesti samanlaisia kuin krokotiilit. Muodostelmasta on löydetty seuraavien choristodereiden jäännöksiä:
- Champsosaurus (vähintään 3 lajia)
- Cteniogenys
Krokotiilit
Muodostelmasta on löydetty seuraavien krokodyylien jäänteitä:
- Albertochampsa
- Leidyosuchus
- vähintään yksi nimetön taksoni
Liskoja
Muodostelmasta on löydetty seuraavien liskojen jäännöksiä:
Plesiosaurukset
Muodostelmasta on löydetty seuraavien plesiosaurusten jäännöksiä:
- Fluvionectes
- määrittelemättömät polykotylidit (lyhyemmät kaulat )
Pterosaurukset
Muodostelmasta on löydetty seuraavien pterosaurusten jäännöksiä:
- Cryodrakon (tunnettu pienistä ja suurista näytteistä)
- 1 nimetön ei-atshdarkidinen pterosaurus
Kilpikonnat
Muodostelmasta on löydetty seuraavien kilpikonnien jäännöksiä:
- Keskity
- " Apalone "
- Aspideretoides (3 lajia)
- Basilemys
- Boremys
- Judithemys
- Neurankylus
- Plesiobaena
- 2 määrittelemätöntä taksonia
Nisäkkäät
Muodostelmasta on löydetty seuraavien nisäkkäiden jäännöksiä:
-
Multituberculata
- Cimexomys sp.
- Cimolodon spp.
- Cimolomys clarki
- Meniscoessus major
- Mesodma primaeva
- nimeämättömät multitubercules
-
Metatherians
- Alphadon halleyi
- Eodelphis browni
- E. cutleri
- 5 lajia " Pediomys "
- Turgidodon russelli
- T. praesagus
-
Eutherilaiset
- Cimolestes sp. (epävarma taksonomia)
- Gypsonictops lewisi
- Paranyctoides sternbergi
- Tuntemattomat therians : vähintään 1 laji
Kalastaa
Muodostelmasta on löydetty seuraavien kalojen jäännöksiä:
-
Chondrichthyans
- Cretorectolobus Olsoni (a partahaikalat )
- Eucrossorhinus microcuspidatus (a partahaikalat )
- Ischyrhiza mira ( sklerorhynchid )
- Meristodonoides montanensis ( hai )
- Myledaphus bipartitus ( säde )
- Protoplatyrhina renae ( kitarakala )
- määrittämätön orectolobid
-
Acipenseriformes ( sammet )
- " Acipenser albertensis "
- Anchiacipenser acanthaspis
- nimetön sampi
- nimettömiä meloja
-
Holostean kala
- Lepisosteus occidentalis ( ga )
- nimetön jousi
- vähintään 2 muuta holosteania
-
Teleost kala
- Belonostomus longirostris
- Cretophareodus ( osteoglossomorph )
- Coriops amnicolus
- Estesox foxi
- Oldmanesox
- Paralbula (mukaan lukien Phyllodus )
- Paratarpon apogerontus ( elopomorf , kuten tarpon )
- vähintään 8 muuta telepostia
Selkärangattomat
Muodostumasta on löydetty seuraavien selkärangattomien jäännöksiä:
- Makean veden simpukat
- Makean veden kotit
- Campeloma (2 lajia)
- Elimia
- Goniobasis (3 lajia)
- Hydrobia
- Lioplacodes (2 lajia)
Kasvisto
Kasvien kehon fossiileja
Muodostelmasta on löydetty seuraavia kasvien kehon fossiileja:
- erilaisia saniaisia
- Equisetum ( Equisetaceae )
-
Gymnosperms
- Platyspiroxylon ( Cupressaceae )
- Podocarpoxylon ( Podocarpaceae )
- Elatocladus ( Taxodiaceae )
- Sequoia (Taxodiaceae)
- Sequoiaxylon (Taxodiaceae)
- Taksidioksyloni (Taxodiaceae)
- Ginkgos
-
Angiospermit
- Artokarpus ( Moraceae )
- Cercidiphyllum ( Cercidiphyllaceae )
- Dombeyopsis ( Sterculiaceae )
- Menispermitit ( Menispermaceae )
- Pistia ( Araceae )
- Platanus ( Platanaceae )
- Vitis ( Vitaceae )
- Trapa ( Trapaceae )
Palynomorfit
Palynomorfit ovat orgaanisia seinämiä sisältäviä mikrofossiileja , kuten itiöt , siitepöly ja levät . Seuraavia palynomorfeja on löydetty kokoonpanosta:
- Tuntemattomat tuottajat
- vähintään 8 lajia
-
Chlorophyta ( vihreät levät ja sinilevät )
- vähintään 12 lajia
-
Pyrrhophyta ( dinoflagellates , eräänlainen merilevä )
- määrittämättömät kystat
-
Bryofyytit ( sammalat , maksat ja sarvikuoret )
-
Anthocerotophyta (sarvikuoret)
- vähintään 5 lajia
-
Marchantiophyta (maksaruohot)
- vähintään 14 lajia
-
Bryophyta (sammal)
- vähintään 5 lajia
-
Anthocerotophyta (sarvikuoret)
-
Lycopodiophyta
-
Lycopodiaceae ( kerhosammat )
- vähintään 11 lajia
-
Selaginellaceae (pienet klubisammat)
- vähintään 6 lajia
-
Isoetaceae ( quillworts )
- vähintään 1 laji
-
Lycopodiaceae ( kerhosammat )
-
Polypodiophyta
-
Osmundaceae ( kaneli saniaiset )
- vähintään 6 lajia
-
Schizaeaceae ( kiipeilevät saniaiset )
- vähintään 20 lajia
-
Gleicheniaceae ( Gleichenia ja liittolaiset; koralli -saniaiset )
- vähintään 5 lajia
-
Cyatheaceae ( Cyathea ja liittolaiset)
- vähintään 4 lajia
-
Dicksoniaceae ( Dicksonia ja liittolaiset)
- vähintään 3 lajia
-
Polypodiaceae ( saniaiset )
- vähintään 4 lajia
-
Matoniaceae
- vähintään 1 laji
-
Marsileaceae
- vähintään 1 laji
-
Osmundaceae ( kaneli saniaiset )
-
Pinophyta ( gymnosperms )
-
Cycadaceae ( sykadit )
- vähintään 3 lajia
-
Caytoniaceae
- vähintään 1 laji
-
Pinaceae ( mäntyjä )
- vähintään 4 lajia
-
Cupressaceae ( sypressit )
- vähintään 3 lajia
-
Podocarpaceae ( Podocarpus ja liittolaiset)
- vähintään 4 lajia
-
Cheirolepidiaceae
- vähintään 2 lajia
-
Ephedraceae ( mormoniteet )
- vähintään 6 lajia
- Tuntematon gymnosperms: vähintään 3 lajia
-
Cycadaceae ( sykadit )
-
Magnoliophyta ( angiosperms )
-
Magnoliopsida ( kaksisikoiset )
-
Buxaceae ( puksipuu )
- vähintään 1 laji
-
Gunnerakasvit ( gunneras )
- vähintään 1 laji
-
Salicaceae ( pajut , puuvilla , haiseva haapa )
- vähintään 1 laji
-
Droseraceae ( auringonkukka )
- vähintään 1 laji
-
Olacaceae ( talipuu )
- vähintään 2 lajia
-
Loranthaceae ( näyttävät mistelit )
- vähintään 1 laji
-
Sapindaceae ( saippuamarja )
- vähintään 1 laji
-
Aceraceae ( vaahterat )
- vähintään 1 laji
-
Proteaceae ( proteas )
- vähintään 9 lajia
-
Komposiitit ( auringonkukat )
- vähintään 1 laji
-
Fagaceae ( pyökit , tammet , kastanjat )
- vähintään 2 lajia
-
Betulaceae ( koivut , leppä )
- vähintään 1 laji
-
Ulmaceae ( jalat )
- vähintään 1 laji
-
Chenopodiaceae ( hanhijalat )
- vähintään 1 laji
-
Buxaceae ( puksipuu )
-
Liliopsida ( yksisirkkaiset )
-
Liliaceae ( liljat )
- vähintään 6 lajia
-
Cyperaceae (sarat)
- vähintään 1 laji
-
Sparganiaceae ( bur-reeds )
- mahdollisesti 1 laji
- Tuntemattomat angiospermit: vähintään 88 lajia
-
Liliaceae ( liljat )
-
Magnoliopsida ( kaksisikoiset )
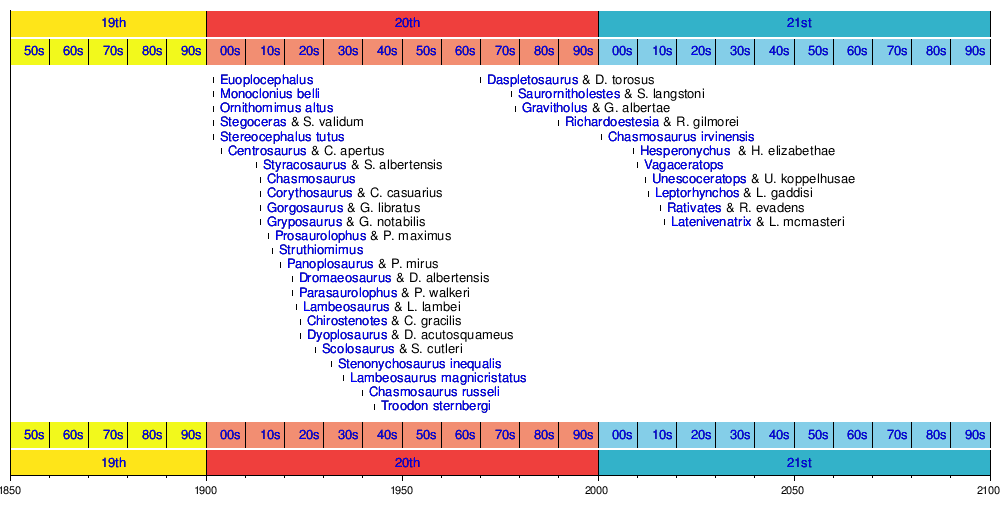
Uusien taksonien aikajana
Seuraava aikajana näyttää kelvolliset taksonit, jotka löydettiin ensin dinosauruksesta. Joitakin lajeja on voitu viitata muihin sukuihin niiden alkuperäisen kuvauksen jälkeen.
Katso myös
Alaviitteet
Viitteet
- Arbor, VM; Burns, ME; Sissons, RL (2009). " Dyoplosaurus acutosquameus -puistojen ankylosaurid -dinosaurusten uudelleenkuvaus , 1924 (Ornithischia: Ankylosauria) ja suvun tarkistus". Lehti selkärangattomien paleontologiasta . 29 (4): 1117–1135. doi : 10.1671/039.029.0405 . S2CID 85665879 .
- Braman, DR, ja Koppelhus, EB 2005. Campanian palynomorphs. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 101-130.
- Brinkman, DB 2005. Kilpikonnat: monimuotoisuus, paleoekologia ja jakelu. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 202-220.
- Caldwell, MW Squatates: alkuperä, filogeenia ja paleoekologia. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.). 2005. '' Dinosaur Provincial Park: Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 235-248.
- Currie, PJ 2005. Teropodit, mukaan lukien linnut. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 367-397.
- Currie, PJ, ja Koppelhus, EB (toim.). 2005. Dinosaur Provincial Park: Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 648 Sivumäärä
- Eberth, DA 2005. Geologia. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 54-82.
- Fox, RC 2005. Myöhäinen liitukauden nisäkkäät. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 417-435.
- K. Gao ja Brinkman, DB 2005. Choristoderes puistosta ja sen läheisyydestä. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 221-234.
- Gardner, JD 2005. Lissamphibians. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 186-201.
- Godfrey, SJ, ja Currie, PJ 2005. Pterosaurs. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 292-311.
- Johnston, PA ja Hendy, AJW 2005. Upper Cretaceous Belly River Groupin nilviäisten paleoekologia. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 139-166.
- Koppelhus, EB 2005. Paleobotanikka. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 131-138.
- Kanadan geologisten yksiköiden sanasto. "Dinosauruspuiston muodostus" . Arkistoitu alkuperäisestä 21.02.2013 . Haettu 29.3.2011 .
- Neuman, AG ja Brinkman, DB 2005. Fluvial -vuoteiden kalat. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 167-185.
- Ryan, MJ ja Evans, DC 2005. Ornithischian dinosaurukset. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 312-348.
- Sato, T., Eberth, DA, Nicholls, EL ja Manabe, M. 2005. Plesiosaurian jäännökset ei-merellisistä rinnakkaissedimentteihin. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 249-276.
- Tanke, DH ja Brett-Surman, MK 2001. Todisteita hautomisesta ja pesimäkokoisista Hadrosauruksista (Reptilia: Ornithischia) Dinosaur Provincial Parkista (Dinosaur Park Formation: Campanian), Alberta, Kanada. s. 206–218. Julkaisussa: Mesozoic selkärankaisten elämä - uusi tutkimus, joka on innoittamana Philip J. Currien paleontologiasta. Toimittanut DH Tanke ja K.Carpenter. Indiana University Press: Bloomington. xviii + 577 s.
- Xiao-Chun Wu. 2005. Crocodylians. Julkaisussa: Currie, PJ, ja Koppelhus, EB (toim.), Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Indiana University Press: Bloomington ja Indianapolis, 277-291