
Epiteelisolu - Epithelioid cell

Yleisen näkökulman mukaan epiteelisolut (kutsutaan myös epitelioidihistosyyteiksi ) ovat aktivoitujen makrofagien johdannaisia, jotka muistuttavat epiteelisoluja .
Rakenne ja toiminta

Rakenteellisesti epithelioidisolut (kun niitä tutkitaan valomikroskoopilla hematoksyliinillä ja eosiinilla värjätyn jälkeen) ovat pitkänomaisia, hienoilla rakeisilla, vaalealla eosinofiilisellä (vaaleanpunaisella) sytoplasmalla ja keskeisillä, munanmuotoisilla ytimillä (soikeat tai pitkänomaiset), jotka ovat vähemmän tiheitä kuin lymfosyyttien . Niillä on epäselvä muoto ja ne näyttävät usein sulautuvan toisiinsa muodostaen aggregaatteja, joita kutsutaan jättimäisiksi soluiksi. Kun tutkittiin transmissioelektronimikroskoopilla sisään Epithelioid soluissa alalla Golgin lamellaarisen monimutkainen on teipattu paitsi zonated, mutta myös tyylikäs vesikkelit tiheä keskus, ja myös hyvin monet (yli 100), suuri granulas, joiden läpimitta 340 nm ja finegranular matriisi enemmän valoa kuin makrofagirakeissa, joskus perigranulaarinen halo. ”Näiden solujen näkyvin piirre on valtava Golgi -alue; jopa 6 yksittäistä Golgi cisternae -pinoa voi olla läsnä sekä muutamia harjaksilla päällystettyjä ja lukuisia sileitä rakkuloita. " Epithelioidisoluissa on tiukasti toisiinsa solukalvot vetoketjullisissa matriiseissa, jotka yhdistävät viereiset solut. Nämä solut ovat keskeisiä granuloomien muodostumisessa, johon liittyy monia vakavia sairauksia. Granuloomissa epiteelisolut suorittavat rajaavat toiminnot.
Sytoskeletonin erityispiirteet

On osoitettu, että filamenttien muodostama epitelioidisolun sytoskeleton eroaa merkittävästi makrofagin sytoskeletonista. Filamenttien määrä kasvaa suuresti näissä soluissa, joissa filamentit (90-100 A) ympäröivät sytokeskusta erottuva rengasmainen nippu, joka usein haarautuu sytoplasmaan. Tällaisen sytoskeletonin vuoksi vierekkäisissä epiteelisoluissa on monimutkainen sytoplasminen interdigitaatio. Käyttämällä pikapakastus- ja jäädytyssubstituutiomenetelmiä (nopea jäädytys, tunkeutuva syövytys ja jäädytyssubstituutio) on osoitettu, että granulatiivisen tulehduksen keskipisteeseen muodostuneet organisaatiot kolmiulotteinen etäpesäkerakenne sytoskeleton, joka on muodostunut granulomatoottisen tulehduksen keskelle . tyypilliselle epiteelisolulle ominaista sytoskeletonia kuin aktiivisten ja liikkuvien makrofagien sytoskeletonille. On esitetty, että välifilamenttien tiheitä verkkoja, jotka on sidottu ytimiin, mitokondrioihin ja muihin organelleihin, valvotaan kaikkialla epiteelisolujen sytoplasmassa. Jotkut aktiinisten filamenttien faskyyrit sijoitettiin alla oleviin filopodiumeihin kuin solukalvot. Solukalvojen täsmällinen interdigitaalinen kolminkertaistuminen liitettävien epiteelisolujen välillä osoitettiin selvästi. Digitaaliset filopodiumit tunnistettiin. Tyypillinen merkki epiteelisoluista on niiden yhdistyminen muodostumalla tiukkoihin interdigitaalisiin kolhuihin "salamana", jolla ilmeisesti voi olla merkitystä tiheän kehon raja -alueen muodostumiselle taudinaiheuttajalta epiteelisolun muodostumisen aikana granuloomat.
Immunologinen fenotyyppi ja immunologiset markkerit
Käytettäessä vasta-aineita RFD9-, RFD7- ja HLA-DR-antigeeneille havaittiin, että kaikilla epiteelisoluilla on immunologinen fenotyyppi RFD9+/RFD7-/HLA-DR+. Saatiin sarja monoklonaalisia vasta-aineita IHY-1, IHY-2, IHY-3, joita voidaan käyttää tunnistamaan tarkasti epiteelisolut, jotka ovat muodostuneet granulomatoottisen tulehduksen etiologisesti eri muodoissa. IHY-1-vasta-aine reagoi sarkoidigranuloomien epiteelisolujen kanssa sekä erilaisten granulomatoottisten sairauksien, mukaan lukien tuberkuloosi, epiteelisolujen kanssa. IHY-2- ja IHY-3 l-vasta-aine, monoklonaalinen vasta-aine, reagoivat epiteelisolujen kanssa sarkoidoosissa, mutta eivät tuberkuloosissa .
Lääketieteellinen merkitys
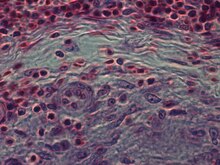
Epiteelisolut ovat olennainen ominaisuus epiteelisolujen granuloomille . Epiteelisolujen granulooma voidaan määritellä spesifisesti ja rakenteellisesti organisoituna epiteelisolujen, makrofagien, lymfosyyttien ja dendriittisolujen kokoelmana. Vieraita kappaleita voidaan pitää järjestäytyneenä makrofagikokoelmana, mukaan lukien pelkästään jättimäisten solujen kokoelmat, jotka ympäröivät inerttejä aineita, kuten ompelumateriaalia-niin kutsuttuja "ei-immuuneja granuloomia". Granulooman muodostuminen liittyy patogeeneihin, jotka ovat oppineet kiertämään isännän immuunijärjestelmää eri tavoin, kuten vastustamalla fagosytoosia ja tappamalla makrofageja. Aineen sulamattomuus makrofagien vaikutuksesta on granulomatoottisen tulehduksen yhteinen piirre. Granuloomat yrittävät murskata nämä organismit ja estää niiden kasvun ja leviämisen. Historiallisesti laajalle levinneet ja tuhoavat sairaudet, kuten tuberkuloosi, lepra ja kuppa, ovat granulomatoottisia tiloja. Granulooman muodostuminen on myös ominaisuus monille nykyaikaisemmille olosuhteille, kuten sieni -infektioille, sarkoidoosille ja Crohnin taudille .
Tieteellisen tutkimuksen historia
Ensimmäinen maininta epithelioidisoluista spesifisenä solumuotona tapahtui 1800-luvulla Koch R: n ja Cornil J : n teoksissa , jotka uskoivat leukosyyttien olevan tuberkuloosin epiteelisolujen alkuperä. Kaneilla tehdyissä kokeissa Yersin A (1888) ja Borrel A (1893) osoittivat, että epiteelisoluja muodostuu veren mononukleaarisista leukosyyteistä. Lewis M (1925) kuvasi ensimmäisen kerran epitelioidisolujen muodostumisen pääkuvioita 1900 -luvun ensimmäisellä puoliskolla. Tämä tutkija osoitti, että veren monosyyttien on soluviljelmissä sekoitettu leukosyyttien Avian (otettu aikuisen linnut sekä alkioiden eri-ikäisiä), hiiressä ja ihmisessä, kun viljelty in vitro , muunnetaan tyypillisiä makrofagien ja Epithelioid soluja, minkä jälkeen muodostamalla jättimäisiä monisoluisia soluja. Lewis M havaitsi epitelioidityyppisten solujen muodostumisen leukosyyttien viljelyn 2.-3. Päivänä. Myöhemmin samankaltaista suunnitelmaa koskevassa tutkimuksessa Jerry S ja Weiss L (1966), kun käytettiin kanan sekaveren leukosyyttien viljelmiä (erotettu Rhode Island Redin sydämen verestä) ja elektronimikroskopiaa, osoittivat, että kanan monosyyttien muutos epiteelisolut alkavat viljelyssä 3-4 päivän kuluttua ja päättyvät 5-6 päivän kuluttua. Koska kaikki aiemmat tutkijat ovat osoittaneet, että epiteelisolut muodostuvat monosyyteistä ja monosyytit ja makrofagit yhdistettiin yhdeksi mononukleaariseksi fagosyyttisysteemiksi , Van Furth et ai. (1972), viitaten Sutton J: n ja Weiss L: n (1966) työhön, antoivat muodollisesti epiteelisolut mononukleaarisille fagosyyttisysteemeille. He eivät kuitenkaan täsmentäneet tarkasti, mistä mononukleaarisen fagosyyttijärjestelmän epiteelisolujen solut ovat peräisin. Samaan aikaan he tekivät erittäin varovaisen johtopäätöksen, että "näissä leesioissa esiintyvät epiteelisolut syntyvät myös monosyyteistä tai makrofageista". Adams D (1976) uskoi, että epitelioidisolut ovat mononukleaarisen fagosyyttijärjestelmän solujen erilaistumisen viimeinen vaihe, muotoili epiteelisolujen sytomorfogeneesin käsitteen, jonka mukaan epiteelisoluja pidetään aktivoitujen makrofagien johdannaisena (joka on useimpien tutkijoiden hallussa tällä hetkellä). Ei perustu oletukseen, että "makrofagien stimulaatio kypsyy edelleen kypsymättömiin epiteelisoluihin ja ultimatelviksi aikuisille". Rhee et ai. (1979) rotilla tehdyissä kokeissa elektronimikroskooppimenetelmää käyttäen osoitti, että yksi epiteelisolujen tärkeimmistä sytomorfologisista piirteistä, jotka erottavat nämä solut makrofageista, on niille ominaisten spesifisten rakeiden läsnäolo, joita he kutsuivat epiteelisolurakeiksi. Omien tietojensa perusteella he tukivat epiteelisolujen sytomorfogeneesin käsitettä, jonka mukaan epiteelisoluja pidetään aktivoitujen makrofagien johdannaisina. : "vesikulaariset" ja "erittävät" epiteelisolut. On ehdotettu, että "vesikulaariset epiteelisolut voivat kehittyä" eritys "-epiteelisoluista degeneraatioprosessin avulla. Etsiessä epiteelisolujen muodostumiseen vaikuttavia immunologisia mekanismeja Cipriano et ai. (2003) saivat tietoja, jotka osoittavat IL- solujen mahdollisen vaikutuksen 4 epitelioidisolujen fenotyypin kaltaisen fenotyypin muodostumisesta makrofageissa.Kaikki kuitenkaan epitelioidisolujen sytomorfogeneesin lakien ja mekanismien tutkimukseen liittyvien tutkimusten tulokset eivät sovi epithelioidisolujen alkuperän käsitteeseen. Deimann J ja Fahimi H (1980) osoittivat, että granuloomien epiteelisolut, jotka on indusoitu rotan maksaan ruiskuttamalla glukaania, beeta-1,30-polyglukoosia, eivät muodostu Kupffer-soluista- kypsistä erilaistuneista makrofageista, vaan verestä De Vos et ai. (1990) saivat tiedot, joiden perusteella he voisivat ehdottaa, että granulomatoottisissa tulehduspisteissä, granulomatoottisissa imusolmuketulehduksissa epiteelisoluja muodostetaan ei erilaistuneesta makrofageja, mutta ns plasmasolumaisten monosyyttien (muistuttavat plasmacytes). Tätä tukevat myös plasmasytoidimonosyyttien ja epiteelisolujen väliset ultrastruktuuriset samankaltaisuudet. Esillä oleva ultrastruktuuraalinen ja immunoelektronimikroskooppinen tutkimus epiteelisolujen granuloomista antoi lisäperusteita tämän hypoteesin hyväksi. Arkhipov S (1997, 2012) käyttäen hiirien peritoneaalisoluviljelmiä, veren leukosyyttejä ja luuydinsoluja osoitti, että makrofagit ja epiteelisolut muodostuvat erityyppisistä monosyyteistä. On osoitettu, että epiteelisoluja muodostuu vain plasmosytoidityyppisistä monosyyteistä, on nimennyt pre-epitelioidisolut ohittaen makrofageiksi erilaistumisen vaiheen . On osoitettu, että kroonisessa tulehduksessa epiteelisolujen erilaistumiseen sitoutuneiden pre-epithelioid-monosyyttisten solujen määrä kasvaa tulehduksen keskellä, veressä ja luuytimessä. käyttämällä hiiren Inbred-linjoja , toisin kuin herkkyyttä Mycobacterium tuberculosisille , osoitettiin, että kroonisessa tulehduksessa muodostuneiden pre-epitelioidisten monosyyttisten solujen määrä on geneettisesti määritetty. Saadut tulokset osoittivat, että epiteelisolujen granuloomien morfogeneesi voidaan määrittää monosytoidityyppisten pre-epitelioidisolujen joukon eri geneettisesti määritellyllä alkutasolla, niiden virtauksella granulomatoottisen tulehduksen keskellä, niiden erilaistumisen voimakkuudeksi epitelioidisoluiksi, ohittamalla makrofageiksi erilaistumisen vaihe ja niiden endomitoottinen aktiivisuus.
Katso myös
Viitteet
Ulkoiset linkit
- Epithelioid+-solut Yhdysvaltain kansallisen lääketieteen kirjaston lääketieteellisten aiheiden otsikoissa (MeSH)
Tämä solubiologian artikkeli on tynkä . Voit auttaa Wikipediaa laajentamalla sitä . |